|
|
新闻资讯
生物物种野生资源调查是其保护的第一步。如果我们对一个物种野外分布在哪儿、还有多少数量都不知道,我们怎么知道它是不是濒危了呢?我们又怎么去保护它们呢?
如果你经常看新闻,或许会看到“XX县发现濒危植物XX的新分布点”之类的报道。如果你偶尔关注一下生物多样性相关的出版物,尤其是介绍各种植物的图鉴,或许你会看到“XX植物分布于XX县”之类的描述。你是否疑惑过:为什么植物学家能知道每种植物分布在哪里?2022年,国家林业和草原局公布的数据显示,我国天然林面积达29.66亿亩。为什么植物学家能在广袤而荒无人烟的山旮旯里找到某一种濒危植物呢? 靠脚? 靠运气? 目前来说,还真是要靠脚、靠运气。 如果你曾经遇到过进山考察的科考队,你会发现他们大多数都灰头土脸、一身泥巴。是的,他们就是那些靠脚去寻找和验证物种野生资源状况的科技工作者。国家和地方人民政府一直支持关于生物多样性的调查项目。其中一类是区域调查,如第一次和第二次青藏高原科考项目、各保护区的调查项目、各地区的本底生物资源调查项目,这类项目对某个区域进行全面调查,遇到什么物种都要记录;另一类是针对某类物种的调查,如2017年科技部支持的“中国西南地区极小种群野生植物调查与种质保存”项目,对西南地区200多种(潜在)极小种群野生植物进行调查和种质资源采集保存,调查的每个物种都要想办法找到全部的分布点。这两类调查是相辅相成的,各自都会形成调查报告和标本等记录。当我们想要知道一个物种在哪儿分布,我们就会去查阅资料,根据记录地点去寻找。 网络普及之后,很多植物爱好者在野外偶遇某种植物后,可能会拍照上传到数据库里(如中国植物图像库PPBC),这些照片的拍摄地点也是重要的分布记录(所以,植物爱好者们!你们真的很重要!)。当然啦,如果保护区护林员在日常巡护中发现了珍稀濒危植物的新分布点,他们也会慷慨地同我们分享信息。有时候,当我们非常迫切地想要知道某种植物的分布点,但又找不到资料的时候,我们会到处向其他研究人员、保护区工作人员及当地群众打听,甚至在已知分布点附近开展地毯式的搜索,比如说我们2016年对极小种群野生植物漾濞槭Acer yangbiense调查时便是如此。但这种方式是需要运气的,非常耗时间,无法覆盖较大的范围,也就没法发现稍远处的分布点。我们在漾濞县蹲守了近1个月,才把漾濞县内的漾濞槭野生资源情况弄清楚。 中国有4000余种受威胁植物(极危CR、濒危EN和易危VU)、1200余种国家重点保护野生植物、纳入国家和地方政府保护规划的280余种极小种群野植物等,不可能每种都用这种方式慢慢地调查。靠“偶遇”更是非常不把稳,大树还好,很多小草本要怎么去发现新分布点呢?比如说兰科铠兰属Corybas 植物(我们公众号介绍过一种可爱的“小红帽”——大理铠兰C. taliensis,感兴趣的读者可以去翻翻),它们高仅几厘米,一株仅1片叶子、1朵花,叶子和花的直径都只有1cm左右,生在深山,就像大海捞针,这要跑废多少腿、花多少钱才能把野生分布点找全呀! (希望读者老爷们读得开心,本文有点长,如果看不懂也没关系(可以直接跳到文末看结论),请理解一下理科生的表达能力,读者老爷们可以看个热闹嘛!别忘了收藏和转发哦!) 野外考察的黑科技 众所周知,每种生物都有它们适合生长的地方,王莲Victoria amazonica长不到高山上,雪莲Saussurea involucrata长不到雨林里,沙漠里也不可能捡到活孔雀。只有在合适的环境中,物种才能自然地生长和繁殖,我们把每个物种对生态环境的需求称为“生态位”(niche)。生态位涵盖的内容非常多,不仅包括气温、降水、光照、土壤等非生物的环境因子,还包括食物链、共生物种、群落等生物因子(看不见的微生物也是群落的重要组成部分哦)。有些物种还具有“特殊癖好”,如依赖火灾,那么火对它们来说也是生态位的重要组成部分。一般来说,我们认为同一个群落中不能存在两个生态位一模一样的物种,一旦存在,它们就会产生竞争。如果一种植物喜欢生活在水边、偏好酸一点的土壤、喜欢强一点的光照,不出意料的话,你可以在很多开阔河谷里找到它们,此时恰好群落里也有另一种植物喜欢这样的环境,那这两种植物就需要去竞争生态位了。值得说明的是,植物可能并不会占据它们所有的理论生态位。举个例子,漾濞槭理论上喜欢生长在森林里某个海拔范围内的水沟边,但是漾濞县合适的水沟只有不到1/3能找到漾濞槭,它们可能争不过同样生态位的植物,所以只能黯然退出群落,也可能只是运气不好,还没来得及把种子扩散到合适生态位。 通过第一轮调查,我们可以收集一些物种分布点的数据,只要把这些分布点上的气候、土壤、群落信息汇总一下,找到规律,不就可以对物种的潜在分布范围进行筛选?对的,这就是“物种分布模型”(species distribution model,SDM)或者“生态位模型”(ecological niche model,ENM)。这是生态学家想出来的“黑科技”,不仅可以用来指导野外调查,还能用来判断入侵物种的入侵范围、计算物种在未来气候环境下的分布范围缩减扩张情况、农作物的适宜栽培区、珍稀濒危物种的就地和迁地保护地规划、推测物种历史上的扩散和进化路径等等。 事实上,“物种分布模型”或“生态位模型”使用非常广。1983-2023年,仅Web of Science数据库里就收录了4914篇直接以"species distribution model"或"ecological niche model"为主题的论文。一般来说,通常将“物种分布模型”和“生态位模型”等同来看,两者使用的建模方法也差不多,但是物种分布不仅仅是由生态位决定的,还受到很多其他因素的影响。比如我们前面提到的扩散能力等,而我们通常仅仅是对其理论生态位进行模拟,所以下文我们都统一使用“生态位模型”(ENM)一词。 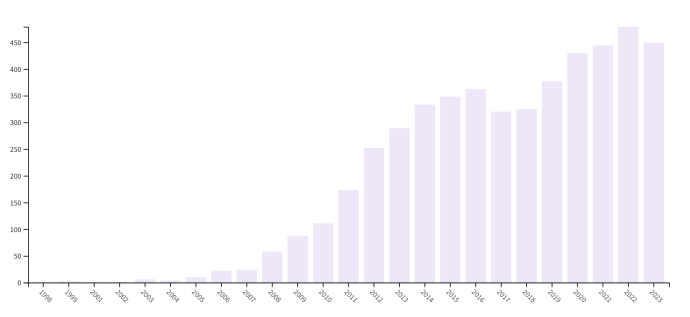 图1. Web of Science数据库里收录的1983年-2023年间主题为"species distribution model"或"ecological niche model"的论文发表数量 2006-2011年,生态位模型的研究突然多了起来。2006年发生了什么呢?其实是MaxEnt模型(最大熵模型)发布了。这个模型还是迄今最常用、也是最方便使用的生态位建模方法,是生态位建模领域引用量最高的模型,引用次数超过1.3万次。2011年,最大熵模型开发团队还为生态学家发表了一篇文章,贴心地告诉大家要怎么使用和解读这个模型。MaxEnt最大的优势是:简单、运算快、建模效果好。它是用Java写的一个软件包,封装得很好,有窗口界面,使用起来很方便,参数也不多,可以用R语言来调用和调参(如通过dismo等很多R包),而且MaxEnt有很好的防过拟合机制,就算全都使用默认参数,建效果也能过得去。  图2. 物种分布模型/生态位模型领域引用最高的文章 MaxEnt是真正的“仅存在点建模方法”。简单来说,假设你是一个模型,我们只告诉你哪些地方有分布点,而不能确定哪些地方没有分布点,那你只需要简单地告诉我:你去找吧,地球上每个角落都可能有这个物种。对呀,你说得没错,甚至正确率是100%!但是这对我们来说有什么用呢?而MaxEnt不会这样去思考,它只需要知道“哪些地方有分布”就能建出很棒的模型了。MaxEnt原理理解起来或许有点困难,但是使用起来非常容易——只要把分布点数据和环境因子地图输入进去就可以,模型会自动输出一个适生度预测地图,还附带各种你需要结果,包括环境因子的重要性、模型效果评价等,无怪乎启发了一大堆相关研究。 MaxEnt也不是全能的,对于稀有物种而言,它还是有比较大的过拟合风险。这也不是它的错,原理很简单:数据量越大,模型越准确,稀有物种一般只有几个分布点,信息量太少了,但是却要找到这几条数据同几十个环境因子之间的关系,这有点强“模”所难。因此,2015年有人开发了一套专门针对稀有物种的建模方法。每次建模只用2个环境因子,大大降低了对数据量的需求,所有环境因子都两两建一个模型,建出一大堆小模型,最后根据这些小模型的评分,把它们的结果合并起来,这种方法被称为“小模型集合”(ensembles of small models,ESMs),对稀有物种有奇效。 最近我们用MaxEnt和ESM对中国的几种铠兰属植物进行了潜在分布区预测,让“小红帽”们也尝尝“黑科技”的力量。 铠兰属植物的生态位分析
大理铠兰Corybas taliensis 呵呵,并不。大理铠兰一般生长在苔藓上,地上部分仅2cm左右,一株有1朵花、1片叶子,花和叶子直径都只有0.6-1cm左右,每年6-7月份开花,其他月份要么只有一片叶子,要么干脆什么也看不到,在地下休眠。哪怕你从一群开着花的大理铠兰身旁走过,你未必能注意到这些小圆点似的大理铠兰。 更何况这些小兰花还专挑人难去的地方生长,比如说喜马拉雅和高黎贡山的腹地,这是人蹲几个月就能调查的吗?中国的几种铠兰属植物除了喜马拉雅铠兰之外几乎都是受威胁的,其中大理铠兰甚至是国家二级保护野生植物。你们知道啥是“珍稀濒危”和保护植物的含金量吗?要是遍地都是、一找一个准,还能叫珍稀濒危植物吗?我再说一遍:植物学家的命也是命!植物学研究生的命也是命!请关爱我们的腿和眼睛!这就是为啥我们不得不对铠兰属使用“黑科技”。 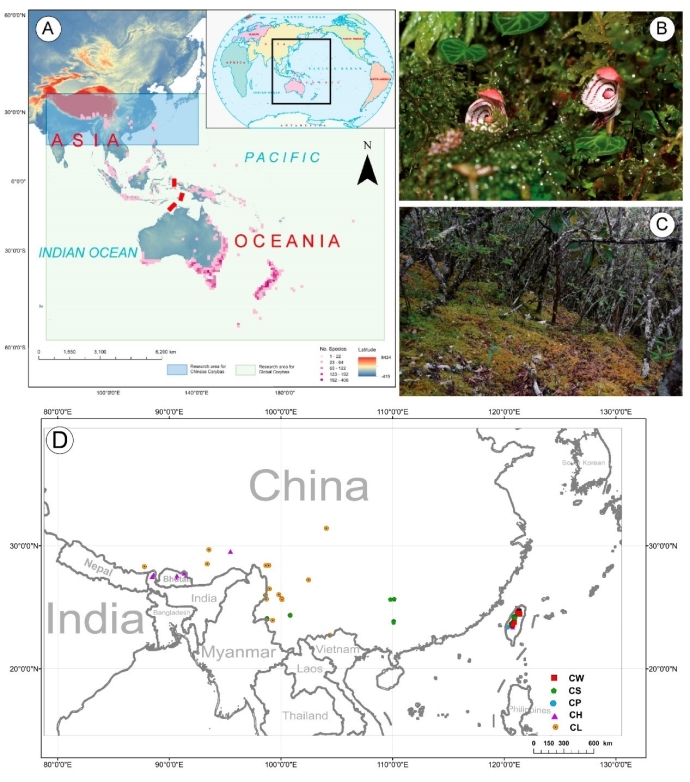 图3. 全球铠兰属分布(A)、大理铠兰(B)、大理铠兰典型生境(C),以及这次研究中使用的5种中国铠兰属植物(D)。CW:台湾铠兰C. taiwanensis、CS:铠兰C. sinii、CP:艳紫铠兰C. puniceus、CH:喜马拉雅铠兰C. himalaicus、CL:大理铠兰C. taliensis。 附注:梵净山铠兰C. fanjingshanensis和杉林溪铠兰C. shanlinshiensis确切分布点较少,没有放入;双凸铠兰C. geminigibbus为中国新分布,只有一个分布点,没有放入。 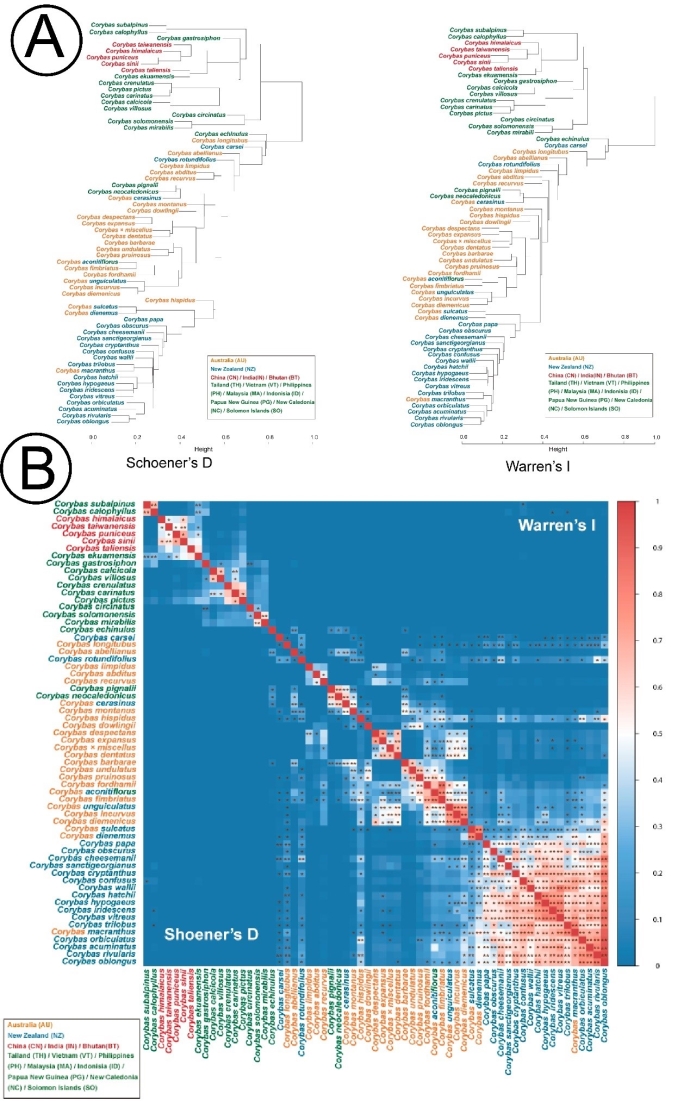 图4. 全球铠兰属植物的生态位距离聚类结果。 附注:中国的物种用红色标出来了,东南亚和太平洋岛屿分布的物种用绿色标出(这个区域很大,但是分布记录太少了,只能把它们混在一起),澳大利亚的物种是橙色,新西兰的物种是蓝色,是不是很有规律?中国的5种铠兰生态位聚在一起,新西兰的铠兰也差不多聚在一起。我是怎么做这个聚类的呢?有个指数叫做“生态位重叠”(niche overlapping),表征的是两个物种之间的生态位相似程度,常用的生态位重叠指数计算方法有Shoener’s D和Warren’s I。我用1减去生态位重叠指数,那不就是两个物种之间的生态位差异(距离)了吗?有距离矩阵不就能聚类了吗?嘿嘿,是不是很聪明。 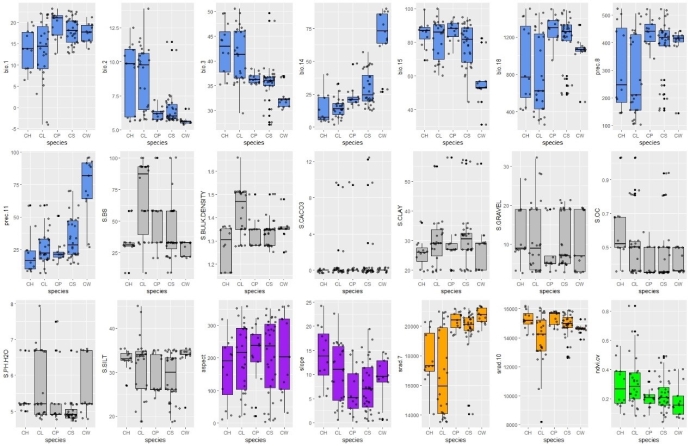 图5. 中国5种铠兰属植物各环境因子的箱线图。蓝色的是气候因子(温度和降水),灰色的土壤因子,紫色的是地形因子,橙色的是光照因子,绿色的是植被因子。 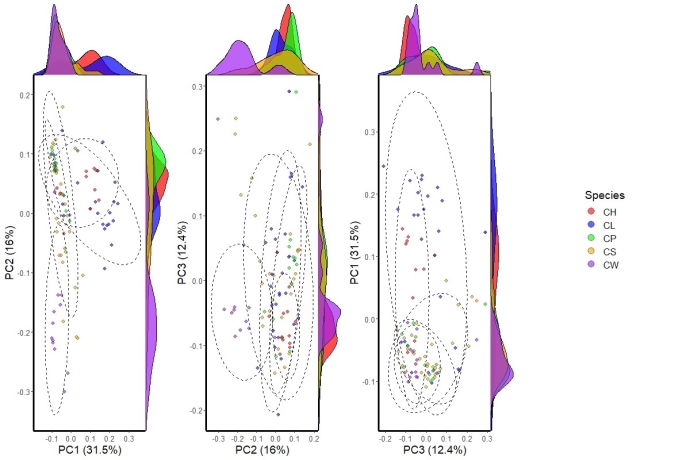 图6. 中国5种铠兰属植物生态位的奇异值分解(SGD)结果。PC1和PC2主要是由气候因子组成,PC3主要是由土壤和地形因子组成。 接下来是我们的重头戏。还记得上文我们介绍了两种生态位模型(MaxEnt和ESMs)吗?我们使用了这两种方法。正巧我们还辛苦地对大理铠兰进行了样方调查,干脆把样方调查的结果也用上,把伴生种当作环境因子之一,看看加入伴生种能不能提高生态位模型的预测能力。建完模型之后,理论上就可以知道当前环境条件下,中国的5种铠兰在哪里比较容易找到。同时我们还可以把模型的结果推广到未来的气候环境下,这样就能预测未来(2081-2100年)这几种铠兰属植物的潜在分布区有什么变化。 您猜怎么着,我们的预测结果居然和前面的中国铠兰属植物生态位差异分析结果十分相似!喜马拉雅铠兰和大理铠兰的潜在分布范围都在青藏高原和云南大部分地区以及四川省的部分地区,对应的气候环境刚好就是干湿分明、植被较密的地区,且喜马拉雅铠兰的潜在分布范围覆盖了大理铠兰的潜在分布范围(图7)。这两个物种形态非常相似,而从生态位的角度看,它们也很相似,因此有可能是同一个物种。因子重要性分析结果显示,喜马拉雅铠兰和大理铠兰对地形、土壤、光照因子比较敏感,而地形、土壤和光照是物种对微环境较敏感的指示因子;但是铠兰、艳紫铠兰和台湾铠兰则不然,它们主要是对气温和降水比较敏感,预测主要分布区在台湾和华南等地。这些预测结果与上面的生态位差异分析结果吻合(图9)。此外,在环境因子已经很充足的情况下,伴生种加入只能对模型效果有微弱提升,但是加入之后可能会取代部分其他因子的预测功能,这说明模型中环境因子的预测能力可能会有饱和,群落伴生种可能是环境因子的一个指针。 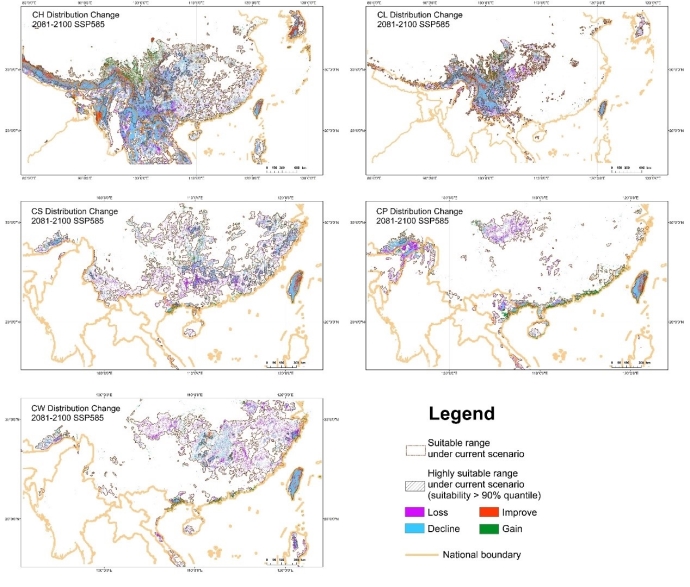 图7. ESM预测的结果 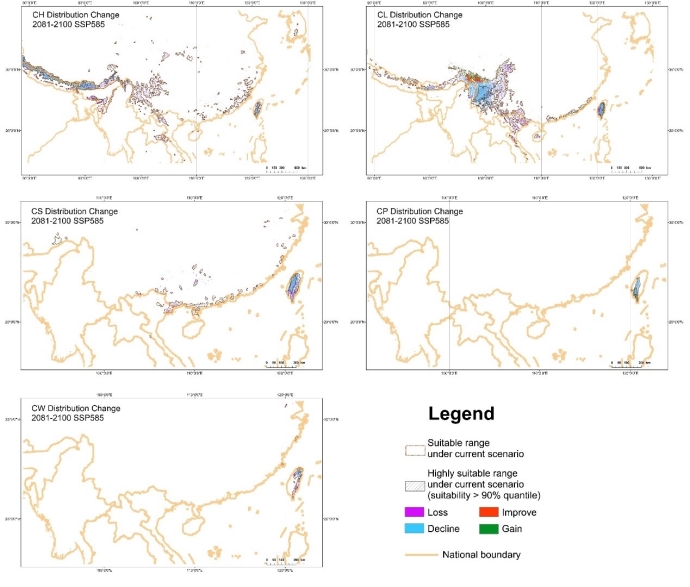 图8. MaxEnt预测的结果  图9. 因子重要性,其中CLBS是加了伴生种的大理铠兰生态位模型。可见,喜马拉雅铠兰和大理铠兰对地形、土壤、光照因子比较敏感,但是铠兰、艳紫铠兰和台湾铠兰则不然,它们主要是对气温和降水比较敏感。另外,伴生种加入之后可能会取代部分其他因子的预测功能。 ESM和MaxEnt的模型评分都非常高,结果大体上比较相似。但MaxEnt看上去有较高的过拟合风险,明明有分布的地方被判定成了不适合分布,可能还是ESM比较适合稀有物种(图7、图8)。 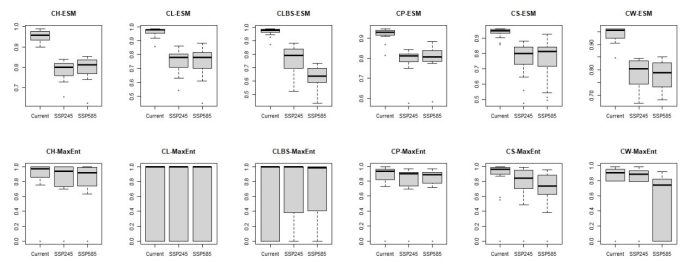 图10. 现存分布点在2081-2100年的适生度变化,可见所有物种现存的居群未来都有显著的适生度下降。CLBS:加了伴生种的大理铠兰分布点适生度预测。 最后,回答大家最关心的问题,我们要优先去哪里找还没发现的铠兰属新分布点呢? 喜马拉雅铠兰:察隅、高黎贡山北部、尼泊尔。 大理铠兰:察隅、川西高原西南部、云南西北部。 铠兰、台湾铠兰和艳紫铠兰:台湾、华南、云南西部、墨脱。 (https://doi.org/10.1016/j.gecco.2024.e03008)。 参考文献: Breiner, F.T., Guisan, A., Bergamini, A., Nobis, M.P., 2015. Overcoming limitations of modelling rare species by using ensembles of small models. Methods Ecol. Evol. 6, 1210-1218. 10.1111/2041-210x.12403. Elith, J., Phillips, S.J., Hastie, T., Dudík, M., Chee, Y.E., Yates, C.J., 2011. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 17(1), 43-57. 10.1111/j.1472-4642.2010.00725.x. Lyon, S.P., 2014. Molecular systematics, biogeography, and mycorrhizal associations in the Acianthinae (Orchidaceae), with a focus on the genus Corybas. University of Wisconsin, Madison. Phillips, S.J., Anderson, R.P., Schapire, R.E., 2006. Maximum entropy modeling of species geographic distributions. Ecol. Model. 190, 231-259. 10.1016/j.ecolmodel.2005.03.026. Tao, L.D., Liu, Y.H., Dao, Z.L., Liu, D.T., Yang, J., Sun, W.B., 2024. Guiding Conservation strategies for China's Corybas species through species distribution modeling. Glob. Ecol. Conserv. 10.1016/j.gecco.2024.e03008. |

版权所有 Copyright © 2002-2016 中科院昆明植物研究所,All Rights Reserved 【滇ICP备05000394号】
地址:中国云南省昆明市蓝黑路132号 邮政编码:650201
点击这里联系我们

